The XXIV Simpósio de Mirmecologia (Myrmecology Symposium) is underway in Belo Horizonte in the beautiful state of Minas Gerais, Brazil! Our editor-in-chief, Benjamin Blanchard, is on site, and The Daily Ant is pleased to cover the activities at this symposium by providing live updates (when Wi-Fi availability is permitting). Enjoy!
Updated at 8:55am on October 9th, 2019
October 4th, 2019
[NOTE: The conference concluded with a final talk by the last keynote speaker, Dr. Mike Kaspari, followed by a closing ceremony. Unfortunately, we had to depart from the conference by 4pm and thus were unable to attend what was surely a vibrant ending to this antsy conference. See here for the official photo of XXIV Simpósio de Mirmecologia. We can’t wait for Mirmeco 2021!]
3:18pm: For the final panel talk of today and the conference, we have have Dr. José Eduardo Serrão on “Trends and Perspectives in ant morphology in the omics era”. Dr. Eduardo Serrão states that “omics” sciences are characterized by their comprehensive unbiased goals (do it all), as well as high-throughput approaches (do it in a single experiment). Several experiments presented provide examples of associations between morphological traits (e.g. glands) and behavior which serve as a reminder that typically, researchers are not interested in morphological variation just to identify variation, but to better understand how animals interact, evolve, etc. Dr. Eduardo Serrão and colleagues have worked on a number of experiments with cross-sections that elucidate in fine detail the feature of ant morphology associated with a number of responses like immune response.



2:46pm: Over in ICB Auditório Cerrado, Dr. Marcio R. Pie is presenting on “Trends and Perspectives in large-scale ant phylogenetics: towards a complete ant tree of life.” For this talk, Dr. Pie will give an overview of common phylogenetic markers, examples of phylogenetic inference, and lastly, efforts to infer a complete tree of life for the ants. The Global Ant Genomics Alliance is working towards getting a full genome for many species, but there are some disadvantages to this approach (relatively expensive, assembly is not always trivial (and neither is annotation!!!), and for many applications it is not necessary to have a full genome. Dr. Pie reviewed (including associated pros and cons) the RADseq approach and sequence capture methods (e.g. UCEs), and also discussed problems with inference like the “anomaly zone” arising from gene discordance/incomplete lineage sorting. To date, no ant phylogenetic inference has included the coalescent model meant to address gene discordance issues. How to get to a full ant phylogeny? One method, used for birds, is to start with a phylogenetic backbone, and attach species under a Yule model based on taxonomic structure. Dr. Pie and colleagues did this for ants: Currently we have 13,599 species recognized by AntCat and 2,640 species with sequence data, and used a backbone from Branstetter et al. (2017). But, we also may not need the “true tree”, and can use e.g. 100’s of trees to account for phylogenetic uncertainty. Oftentimes downstream analyses (e.g. trait evolution) are robust even with phylogenetic uncertainty incorporated into the analyses.


2:21pm: Now, Dr. Heraldo Vasconcelos on “Trends and perspectives from long-term data on ant communities”. Studies on the temporal dynamics in ants are rare, due to various difficulties (like the timeline that graduate students have to conduct work!). The most common approach is the “chronosequence approach”, utilizing a “space-for-time” substitution. Sometimes there can be not enough replicates, lack of interspersion among treatment plots, and pre-existing differences across plots. If you want to know how “undisturbed” communities change over time, or how communities respond to (“subtle”) disturbances, this approach is often insufficient. Long-term monitoring of the same, unmanipulated plots can be better for such questions, which allows researchers to track natural responses to disturbance. What about ants??? Donoso (2017, Ecological Indicators) is the only paper of this sort to date, sampled same community in Ecuador 6 times during 11 years, found tropical ant communities are in long-term equilibrium. But how are ant species responding to climate change? Longterm monitoring necessary to address this question. But how can we get these data? Establish plots, but need funding and these are limited in spatial coverage. One alternative is re-surveying “old plots” that were inventoried before. Case study: Dr. Heraldo Vasconcelos and colleagues used this approach, with arboreal pitfall trapping over 2005-2019 (two sites with unequal histories of fire disturbance), analyzing trajectories. In another case, they assessed ants associated with an EFN-producing tree pre-fire, resampled after a fire in 2017 and 2018. Thinking more broadly again, resampling old plots can be difficult sometimes, if not marked well or not georefrences – and also seasonal and observer biases as well. Ant-specific problems are variation in sampling methods, bias in ant sorting and identification, and vouchers not deposited or properly labeled. Outstanding issues include how to deal with morphospecies, unifying research collections, establishing collaborative research groups, etc.


2:00pm: After lunch, we’re once again the main auditorium to hear Dr. Heloise Gibb speak on “Trends and perspectives in trait-based approaches to understanding ant community structure”. Dr. Gibb opens discussing the study of functional traits and the different approaches used to get at questions such as the realized and fundamental niche of a species/group. Field studies, manipulative experiments, gradients all important here! Dr. Gibb and colleagues utilize a response-effect framework for ants. Which traits to use? Often focus on morphological traits, especially ones easier to measure, but might be good to expand upon these too. The Global Ant Database, created and maintained by Dr. Gibb and colleagues, is similar in this respect. One use of such databases is studying trait correlations. Dr. Gibb also notes how ants are considered like plants, as noted in Andersen (1995). Comparing assemblages in trait space is difficult, as globally most variation in traits is within community and intraspecifc variation is significant. Dr. Gibb and colleagues have utilized the global ant database to understand functional richness, diversity, evenness; morphological diversity; and other response variables. For a bit more on the Global Ant Database: An open-access repository with locality, assemblage, and traits data, as well as a traits guide. Would like to link with other ant databases as well as other taxon trait databases. We need, as a field, further work with such data, including identifying trait syndromes/trade-offs, mechanisms, accounting for intraspecific variation, 3D data, etc.




12:10-2:00pm: Almoço delicioso!
12:10pm: A snippet of summarizing thoughts by Dr. Leal:
12:02pm: Dr. Leal additionally presented work with colleagues on Atta leaf-cutter ants, showing that leaf-cutting populations benefit from human disturbances in the Caatinga, as well as assessing impacts on soil quality, showing that soil fertility is higher in refuse dumps and is not impacts by disturbance or aridity (among other results). Furthermore, Atta nests appear to be seed sinks in the region (as determined by extensive colony excavation).



11:53am: Next, Dr. Leal shares that the interaction between ants and EFN-bearing plants is very conspicuous in Caatinga. EFNs appear to structure ant communities in arboreal communities in the Caatinga. They measured the stability of ant-plant networks to disturbance, and found that specialization decreases with disturbance and aridity increases. Because of the increase in generalist ant-EFN plant interaction, the attack of herbivores is reduced as rainfall decreases (although chronic disturbance did not affect these services).





11:46am: Ant-plant interactions! Dr. Leal and colleagues have recorded that Caatinga is a hot spot for myrmecochory (seed-harvesting and dispersal by ants). Through a number of studies, they found that removal rate and distance decrease as chronic disturbance increases, but only for high-quality disperser ants and for preferred diaspores; that ant-diaspore interaction, removal rate, and distance decrease as aridity increases, but is not affected by chronic disturbance; and that Dinoponera nests present higher seedling richness and densities compared to control areas, but again no relationship with disturbance. Overall, changes in seed dispersal to disturbance are influenced by ant partner abundance and behavior.
11:32am: Now, what about ant response to chronic anthropogenic disturbance? Dr. Leal utilizes the ant functional group classification used by Alan Andersen. Both opportunists and specialist predators negatively impacted by disturbance. In a longterm ecological research project, Dr. Leal and colleagues assess impact from differential rainfall and human land use on ants. See Arnan et al. (2018, Ecological Indicators) for more! Dr. Leal then went quickly through results from several other studies conducted with students/collaborators – see the below pictures for some of these results.





11:20am: We have a second plenary session this morning, by keynote speaker Dr. Inara R. Leal Caatinga on “Global changes and the future of Caatinga dry forest: lessons from ants”. They open mentioning, in humor, that Caatinga is the “ugly duck” of ecosystems in Brazil. Also, much of the presented work is reported in a recent book on the Caatinga ecosystem of Brazil! Many areas are under intense pressure from human activity. What is the Caatinga? A dry forest habitat that is a mosaic of seasonally dry tropical forest and scrub vegetation and makes up 10% of Brazil. ~28 million people which depend on Caatinga resources for their livelihood – firewood collection is more than 600 kg/person/year (!) among other timber and non-timber product usage. And, Dr. Leal notes that this region is being impacted by climate change as well. What do we know about ants in the Caatinga? Not much, actually! Only paper at regional scale is in Ulysséa & Brandão (2013). Leal et al. (2017) added species collected from recent surveys; total species number is 276 species, 62 genera, and 10 subfamilies. And the species accumulation curve suggest that many species are yet to be discovered. Dr. Leal tells us that, perhaps surprisingly, only a couple known endemic ant species are known from the Caatinga – the fauna appears to largely be a subset of the Cerrado fauna.



11:00am: To close out this morning open talk session, Jignasha Rani is telling us about her work on “Drivers of Diversification in a widespread species complex within the continental adaptive radiation of the turtle ants (Cephalotes)”. Turtle ants are a neotropical “success story”, and also a good system to investigate less-studied “cryptic” diversification. The wide distribution of C. maculatus is uncharacteristic of an adaptive radiation, where species tend to exhibit narrower distributions, and the worker caste is relatively uniform in morphology while the soldiers are less well known. Rani proposes three different hypotheses: The species is a geographically widespread species, the species is undergoing speciation due to geographic isolation, or the “species” is actually a complex of differentiated species. Utilizing UCE data, Rana and colleagues found little differentiation, with specimens spanning from Costa Rica to northern Argentina, supporting the widespread hypothesis. However, there actually is some structure in the C. maculatus clade, with Trinidad specimens and a few specimens from Minas Gerais (!) differentiated some from all the others (some sympatric with the differentiated specimens in Minas Gerais). To further explore the potential that this species is an adaptive differentiation, Rana and colleagues look at morphological differentiation in the soldier caste of C. maculatus. They found some apparent variation, including head with, posterior margin of the head, and hair differences. So, there is potential for an adaptive radiation here, but there is more to come both for the morphological analyses as well as population genetics work.


10:42am: Shifting to morphology, we have Dr. Paco Hita Garcia (again!) presenting “Morphology reloaded (again) – ANTSCAN or how to connect phenotypes with genotypes through next-generation phenomics”. Some slides for this talk are similar to the one yesterday (with apologies by Dr. Hita Garcia) [Note: So we won’t copy that information here]. There is a pronounced disparity in amounts of molecular vs. morphological data, as well as a large gap in scalability, but modern tools and technology should be able to bridge this gap (and problems with things like data size/storage is getting better and better). So what is ANTSCAN? Wants to “Scan all ants”, provide full phenomic data for the same and many more ant genera as used in the GAGA (global ant genome) project. The pipeline used by Dr. Hita Garcia and colleagues is “near perfect” for skeletal/cuticular and musculature structures, good but few details for the brain/nervous system, variable for glands, and currently not possible for the tracheal system. A collaborator in Germany has a faster approach with a synchrotron high-throughput x-ray tomography system (“we can scan all ants in literally one week”!). Goal is to get three days of this system and scan ca. 2000 specimens, or at least as much as possible (sub-projects in Japan, Switzerland, SE China, and Brazil, as well as an eye size-brain size-ecology project, surely among others). Overall, aim is to cover as much of the available ant diversity as possible and to make data freely available!



10:27am: Next, we have Gabriela Procópio Camacho speaking on “Unraveling diversity patterns and endemism in Malagasy ants: A Phylogenomic approach”. The goals of this project: Describe patterns of species richness, phylogenetic diversity, and endemism, and to identify centers of diversity and endemism important for arthropod conservation. Procópio Camacho and colleagues are using a phylogenomics approach with ~2,000 UCE loci, 1,212 species, and non-destructive methods. They will join this with species distribution data to model species distribution based on climatic niche data as well. Presenting preliminary results for the genus Crematogaster, they find variable overdispersion and clustering patterns. Weighted and phylogenetic endemism metrics reveal similar patterns, but centers of endemism patterns are slightly different. There are zones of “neo-endemism”, “paleo-endemism”, “mixed endemism”, and “super endemism”. The overall centers of endemism are similar for lemurs, reptiles, and amphibians, as well as low overlap with protected areas. Next steps are to conduct similar work across all ants in Madagascar (and define areas that are currently under sampled).



10:12am: After a brief break, we’re still in the main auditorium to hear Rodolfo da Silva Probst talk about their work on “UCE Phylogenomics Reveals the Evolutionary History of the Arboreal Ant Genus Myrmelachista (Formicinae: Myrmelachistini)”. The study genus is an ant-plant mutualist, and the first result presented is that dead-stem specialists evolved twice, and that there is independent evolution of specialist live-stem (utilizing the same plant species!). A dated phylogeny was also presented. Next, Probst presented work where they extracted COI data from UCE data, and utilized IQ-TREE tree for inference. Compared to a COI tree by itself, which is weird in some ways (though overall matched UCE topology), the merged tree revealed some interesting patterns (and some polytomies). Probst then quickly presented a couple of other results, and noted that future work will focus on convergent evolution of specific relationships in Central America.



9:50am: Lessons Dr. Parr learned: (1) Aim big, but start small (pilot studies, proof of concept). (2) Risky ideas often don’t work the first time, but you can learn from them. (3) It is really valuable to form and maintain a network of collaborators. (4) Don’t give up if you think you have a genuinely stellar idea. (5) Don’t be constrained by what you think is feasible – think big first, and then reduce it down to what you can do feasibly (with money etc.). But what if you just don’t have an amazing idea yet? Surround yourself with inspiring, curious, innovative, ambitious, open, and fun individuals! And make time to think! Read! Observe! Get into your study system!
9:45am: There’s lots we still don’t know! This is important to remember, Dr. Parr reminds us. And complex systems can result in surprises for researchers. A study that placed carbohydrate and protein baits along a gradient up a tree found that, unexpectedly, that ant suppression was found even up the tree, despite the fact that the poison baits were only implemented on the ground. But what is the mechanism? Could be time of day (ants coming down to the ground at times not overlapping with trial periods) – alternative is that they could also be being competitively excluded by ants on the ground.
9:41am: Termite time! To determine interactions between ants and termites, Dr. Parr and colleagues added in termite exclusion plots too. Termite soil removal was higher under ant suppression conditions. Decomposition was tested with… toilet paper (highly technical). Next: Herbivory, where we’d expect a higher abundance of herbivores and thus higher rates of leaf damage.



9:34am: Dr. Parr now moves on to consider: What roles might ants be playing? One main role seems to be scavenging. Utilizing cages and uncaged baits, Dr. Parr and colleagues could test the impact of ants and other insects, and suppressed ants to test the specific role of ants. Notably, this is an underestimate (!), as not all ants could be excluded. This experiment was also repeated in South Africa, with very similar ants. One difference was that ants removed somewhat less percentage of the baits (> 30% vs. >50%), but this could be due to differences in available foraging hours as the savannah is much hotter, and removal per hour may be similar.


9:35am: The Myrmecosphere!
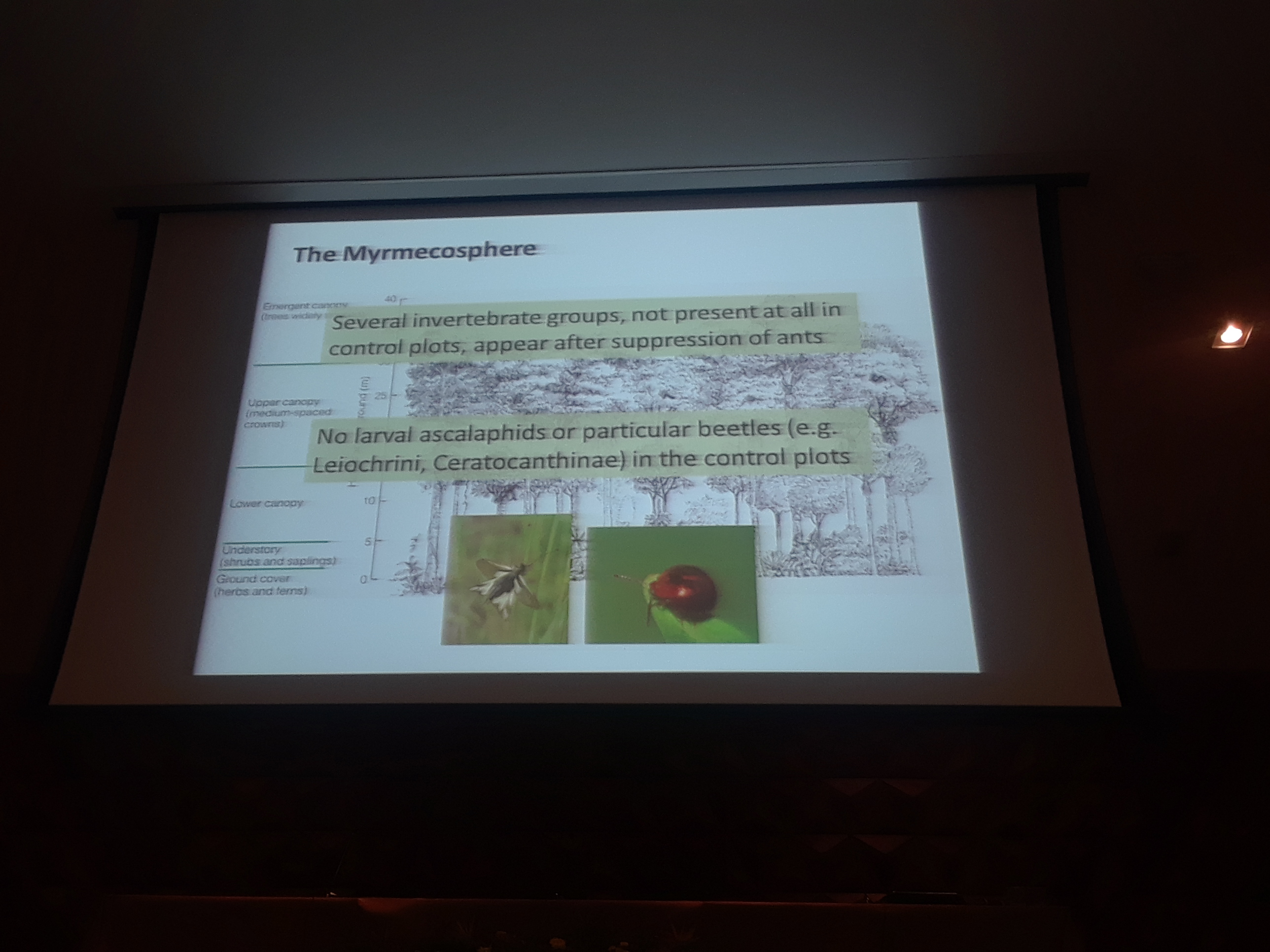
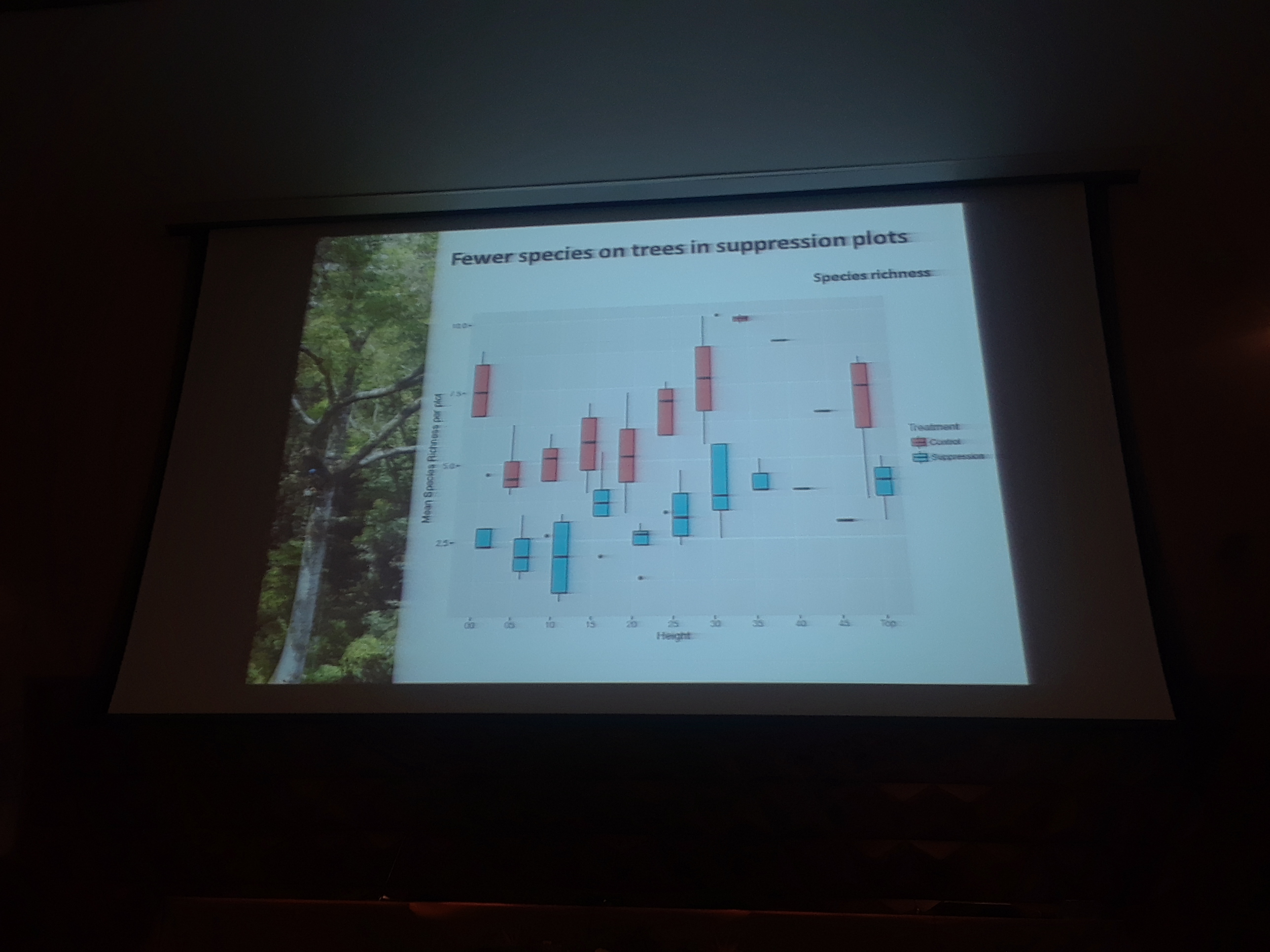
9:20am: Now, to Maliau, Bornea, to try a similar experiment (replication!). A similar set-up was used to that in South Africa. Dr. Parr and colleagues similarly seed, can we suppress the abundance of ants? Do ants structure the invertebrate community? What effect do they have on ecosystem services? To the first question, yes, mean reduction of 93%, and all genera suppressed, similar to the South Africa results. What about structuring of the invert community? In a course-scale analyses, they looked at Winkler leaf litter sampling and identified to the order level. Several common insect groups significantly increased when ants were suppressed. Predators and decomposers do much better as well, and mobility (ability to escape the ants) seems to matter (more mobile groups responded less to ant suppression, and immobile groups as well, likely due to defensive traits already established). Looking just at beetles, it was “a bit of a nightmare” given the diversity of beetles, but most of the groups that increased were predators. A few groups also decreased, and the reason is unsure at the moment. Next, Dr. Parr considered myrmecophiles (ant-loving species that benefit directly from ants), and found that some obligately myrmecophile beetle species are lost in the ant suppression plots, a nice validation of the ant suppression.



9:13am: Big thinking about the little things… some work in ants! Ants and termites are dominant in the tropics – more than 60% of insects in the leaf litter in the tropics are estimated to be ants. “The little things that run the world”, a famous quote by E.O. Wilson, is a hard statement to test. Enter the TARTs (Termite & Ant Research Team), to utilize large-scale projects to address the role of ants in the world. What do ants do at the ecosystem level? Dr. Parr and colleagues set up a large scale ant exclusion/suppression experiment in South Africa, utilizing a poison baits treatment reduce ant numbers. Cat food + jelly + ant poison was used for the bait – yum! Mean reduction of 76% in the plots (here, Dr. Parr apologized for feeling like “Dr. Death” in this ant conference, to audience laughter), and ant genera were equally affected. Did the poison inadvertently impact other invertebrates, though? No, which is good! See Parr et al. (2016, Ecology) for more.


9:04am: Dr. Parr opens on risk-taking and attempting large-scale projects. Such projects can be some assortment of long-term, large-scale, novel, in a difficult system, or utilizing new technology. What sort of ambitious experiments in ecology that have inspired Dr. Parr? One example is Simberloff & Wilson (1968), the “Florida Keys experiment” in the mangrove islands, that involved fumigating small mangrove islands and monitoring the recolonization of the islands (probably not something you could do nowadays!). This work led to core concepts in biogeography. Another example, Paine (1969), where the researcher removed the sea star and demonstrated a “keystone” role. At the time, studies were often more observational, so this was “big science” in the sense of being novel. Other examples include Schindler (1974), and 8-year project on Great Lakes eutrophication; Brown & Davidson (1977), the Portal Project that studies the exclusion of small mammals and impacts on seed harvesting ants [or also excluding seed harvesting ants?] and the biological community – still ongoing today, and a rare example of long-term studies with insects! Why are such studies rare? Expensive, risky, low replication, perceived as less rigorous.


9:00am: All good things come to an end, and alas, this is true even for the most good of things (the ants). Thus, we arrive at the final day of this exciting conference full of a wide variety of ant research! To start off the beginning of the end, today’s morning keynote speaker, Dr. Kate Parr, is about to give a talk on “Thinking Big About the Little Things”!

October 3rd, 2019
5:34pm: In closing, Dr. Eugênia de Carvalho Campos thanks those who worked on the specialization course in urban entomology:
5:18pm: At the start of the third part of the talk, Dr. Eugênia de Carvalho Campos introduced the topic of urban pest control in Brazil by discussing how understanding of ant pests was developed in the mid- and late-1990’s – before this time, knowledge of how to properly control ant pests was very limited. Together with colleagues, they developed a course, a specialization course in urban entomology, to enhance knowledge about urban tramp species (including ants). This program, about 460 hours, covered everything from how to conduct a literature review to ant morphology to toxicology. Participants are also trained how to create a management and control plan. Of 142 students in the course, 27 students chose to work with urban ants! And the reports are freely available online. What has changed, after this course? Now, Brazil has nine registered baits, spraying formulations, 142 specialists, and literature in Portuguese.


5:09pm: Now, for… fire ants! The mitochondrial gene COI was collected from a number of fire ants to document the population biology of the group. In another study, Dr. Eugênia de Carvalho Campos investigated carpenter ants and the impacts on tree fall in cities. Over four years, they surveyed more than 1,300 trees and 138 palm trees, and carpenter ants were associated to the plant species. Chemical control was performed as well. What did they find? 7.6% of the tree species had carpenter ants, mostly Camponotus atriceps. Next, they wanted to conduct a better survey, including the genetic diversity of Camponotus in São Paulo as well as alates. In these collections, they found 16 species of Camponotus, which is a high diversity for a city as big as São Paulo! Richness was higher in less stressed areas (fewer residents), and including alates (assisted by molecular biology) captured several species not sampled through workers. Importantly, using alate is a valuable tool for surveying biodiversity in urban and arboreal habitats.


4:59pm: Further work found that ant species often occupy hospitals, up to nine species in one hospital. Then, at an international scientific meeting in Campo Grande, MS (1989), was focused primarily on Solenopsis invicta and leaf cutting ants. [Note: We missed a transition here]. Dr. Eugênia de Carvalho Campos notes that “ants are everywhere varying in richness and occurrence, depending on the structure of each environment”, and can sting, decrease biodiversity, damage equipment, cause high infestations and generally be a nuisance for humans. These include species that are native ants that are considered invasive elsewhere. Also, it is interesting that ant nuptial flights are really observed in urban areas, while in non-urban areas, there are reports of nuptial flights with large ants – perhaps there are fewer Ponerine alate in urban areas? Dr. Eugênia de Carvalho Campos conducted a study on alates in urban areas documenting the number of alate specimens of Ponerine ants collected with light traps, in two regions in the city of São Paulo, Brazil, from October 2012 to October 2013. Many Ponerine alates were collected, most from one genus [We believe we saw this correctly].
4:46pm: Arriving a bit late to the talk, due to getting caught up in engaging conversation during coffee break, we’re now in the [Main Auditorium] to hear keynote speaker Dr. Ana Eugênia de Carvalho Campos on “Science and classroom, a recipe to control urban ants”. Dr. Eugênia de Carvalho Campos is now speaking about the first formal urban ant records in Brazil – a later paper on these records have noted that tramp ants were not present in those earlier days. Tramp ants have the characteristic of not building a nest but rather occupying cavities and species already existing, e.g. in human homes. Professor Eugênia de Carvalho Campos found a record of the pharaoh ant from 1936, and then searched and found a 1931 letter section to a newspaper that notes “several species in residences in Brazil” that are tramp species (including the Argentine ant, tawny crazy ant, the pharaoh ant, and a couple tramp carpenter ants). In the 70’s, equipment from East Germany was sold to Brazil in UNESP, and the boxes contained pharaoh ants, and occupied the whole building – this was a problem, as they started messing up other invertebrate trials by transferring bacteria to where they were contained, killing them.

3:57pm: For the final talk of this section, Dr. Maurice Leponce speaks on “Three-dimensional distribution of arboreal ant in rainforests and a method to detect ant mosaics”. Evidence of ant mosaics is often indirect, using co-occurance statistics. There is, also, diversity and stratification of the ant assemblage, with the arboreal zone often excluded. Methods to incorporate arboreal ants include tree climbing, insecticide fogging, canopy cranes, and tree felling, all of which have varying pros and cons. The aims of the work presented today are to develop a a rapid assessment protocol to detect dominant ants (and associations between them and 3D colony expanse), evaluate protocol representativeness, and compare behavioral vs. statistical approaches to detect ant mosaics. Two useful methods that have been used by Dr. Leponce and colleagues are baits that are wrapped with paper towel and using a sling shot to place baits along a tree at different heights. Implementing aggression tests has assisted to understand associations between dominant ants. To evaluate the extent of a colony in 3D space, intraspecific aggression tests are useful as well (nestmates will not fight). Dr. Leponce and colleagues documented how different groups of ants (dominant, non-dominant, and ground-nesting) shift as the arboreal strata differs. In evaluating the representativeness of the protocol, about 55% of the species are captured [Note: We may not have fully understood the result, here!]. One thing confounding for statistical analyses is co-occurrences but segregation by strata.



2:58pm: Moving to the Auditório Campo Rupestre, Dr. Stacy Philpott is presenting on “Ants and their ecosystem services in Mexican agroecological systems”. Dr. Philpott reminds us that there is a mutualism between ants and green scale insects in Mexico coffee systems, and also notes that, of course, coffee is economically important! Besides this economic importance, it also carries great environmental importance given that coffee is cultivated in 16 of 34 biodiversity hotspots. Coffee is a crop often grown under trees (either diverse or more sparse), and sometimes under no shade as well. Perhaps unsurprisingly, then, change in coffee management can impact ant communities including richness. There are about 120 species that nest in the ground, 40 that nest in twigs in trees, and 20 other arboreal ant species, in the systems studied by Dr. Philpott. One study by Dr. Philpott and colleagues found that Azteca presence on shade trees and coffee plants affects twig-nest ant colonization and composition. Furthermore, pest species, like the coffee berry borer, can play an important role in these communities as well. Does adding arboreal connections improve predation services provided by ants? What about ant diversity or phorid fly (parasitoid) presence? Seasonality and microclimate? Dr. Philpott and colleagues addressed these questions through a series of studies. In the first, they found, through manipulative experiments, that activity increased in connect plants significantly more than in control plants (when string was added. Furthermore, the number of borers removed was three times higher in connected plants. In a second study, they investigated three specialist phorid fly species, which parasitize Azteca and limit foraging by the ant workers. Using tents with coffee plants a series of insect presence trials, they found that phorid flies significantly reduced the benefit of defense (borer removal) by the ants, when all three groups were present. But after adding one or two additional ant species, the beneficial defense was restored: more ant diversity = more coffee! Now, for the third study, Dr. Philpott and colleagues consider if coffee borers are being predated on on the ground (as they noted that Azteca ants remove borers by throwing them to the ground off the coffee plant). They found very low removal from the ground, by ants, during the rainy season, and much higher during the dry season. Predation was also higher when there were lower temps and higher leaf litter as well. Notably, there were no differences in the ant species identity that removed the borers between seasons. What about prey availability? Most species were more predaceous during the rainy season, consistent with prey availability as a driving factor behind these results. Farmers could utilize these results to boost coffee productivity!



2:26pm: Next, “So long, and thanks for all the features: Is the head shape in Pheidole modular and is it predicted by food preference?”, by Alexandre Casadei Ferreira. Pheidole, also known as big-headed ants, are characterized by polymorphism, with major workers and minor workers. There is a lot of variation in head shape between subcastes, of course, but also within subcastes. To ask how head shape evolves, Casadei Ferreira and colleagues used two landmarks and 20 semi-landmarks defining the outline of the head. They found significant overlap in shape, between majors and minors and also between “old world” and “new world” groups of Pheidole. They also detected a bifurcated diversification of shape for both minors and majors, when incorporating phylogeny. What about mandibular shape? Casadei Ferreira and colleagues also investigate mandibular shape using micro-ct data. The database included 28 species and 56 specimens, representing 5 granivorous species, 14 generalists, and 9 species of unknown diets. When testing to see relations between head shape and nesting type or diet, no differentiated relationship was detected. With the mandible morphospace, similarly, no differentiated relationship was detectable. This finding is similar to a study that found that the shapes of bird beaks are highly controlled by non-dietary factors. For future work, Casadei Ferreira would like to investigate more complex models, for example head + size + diet + nest, to explain the evolution of shape in Pheidole.


2:00pm: The morning was filled with ant outreach, and now the afternoon is filled with ant talks. Here in the Auditório Cerrado, we’re starting off with Dr. Paco Hita Garcia on “Ghost in a shell: Next-generation phenomics and 3D cybertaxonomy in ants”. We have entered the era of big data in biology – the “ics”, thus calling use of 3D imaging to understand traits as “phenomics”. The past decade has seen significant advances in imagine, including x-ray microtomography (micro-ct). Scans can take quite long sometimes, even up to 30 hours for one sample, but as quick as 27 minutes too. Processing these data is quite time-intensive, involving segmenting (and virtually dissecting) the data based on color and contrast on a computer. One way to use these data is producing 3D surface models and, for example, make videos with them. This is a great resource for both science and public outreach! How about use in taxonomy? Yep: Diagnostic characters often traditionally require dissections (e.g. to study male genitalia), but micro-ct can get around this requirement. These techniques may also allow taxonomists to discover and evaluate new diagnostic morphological characters. Dr. Hita Garcia also presents the concept of “cybertype”, a micro-ct scan of the type specimen (the one used to describe the species) saved online, easily retrievable and freely accessible. However, micro-ct scanning does not capture color, so cybertypes should be directly associated with 2-D images of the specimen, in color. After describing this, Dr. Hita Garcia moved to discussion of a project, with colleagues, on diversification in Madagascar ants. They sampled five species-rich Myrmicine genera, and combine phylogenetic inference, geographic data, climate data, and geometric morphometrics using 3D shape data. Can we understand the processes underlying radiations of species-rich ant genera? No results yet, but should be ready by the next meeting! And finally, Dr. Hita Garcia quickly presented some data showing that qualitative morphological data does not explain phylogenetic relationships while 3D shape data confirms geographic signal in the data in Malagasy Terataner ants.



12:00-2:00pm: Almoço delicioso!
11:23am: An ant enthusiast from the local university (UFMG) is also on site with some homemade structures that house ant colonies – wonderful for personal enjoyment and community outreach alike! The kids are engaged and curious.

10:20am: The kids got very excited to answer questions and see some new ants!
10:14am: Dr. Manu Ramalho sharing about ants!
9:48am: This morning, the conference day opens with a wonderful, 3-hour event: “A Tour to an Ant Nest”! What is this event?
By the end of the morning, around 400 children from local schools will have descended upon the conference site to hear about ants and, more importantly, tell us about their experiences with ants! One of the main activities is drawing pictures of ants and things that ants do. It’s already quite a treat to see so many kids excited about formigas!!!
October 2nd, 2019
5:38pm: Now for some work on stability in networks – redundancy is important. Network stability should increase with more species and redundancy, thus higher complexity. Why is this? Differences between “redundant” species and the portfolio effect (diversity driving stability. After some examples including ants, Dr. Blüthgen closed with a brief summary:
5:27pm: Networks can also be used, of course, for ant-ant mutualisms, which exist! They looked at a Crematogaster–Camponotus mutualism, which support each other in a given space, and also recognize their specific mutualistic colony and will be antagonistic to individuals of the same species who are not paired with them. Research showed that chemical signals that structure this interactions, including discovering a new class of chemical, named “Crematoenone” after Crematogaster, maintaining the mutualism. Blüthgen and colleagues also found a similar chemical-based network in stingless bees and for omnivorous ants (via lipid fatty acids).
5:19pm: Army ants are highly specialized in prey ant species… somewhat surprising! Dr. Blüthgen also as worked on ant-plant wound sap networks, common in, for example, southeast China – more on this in Staab et al. (2017, The American Naturalist).
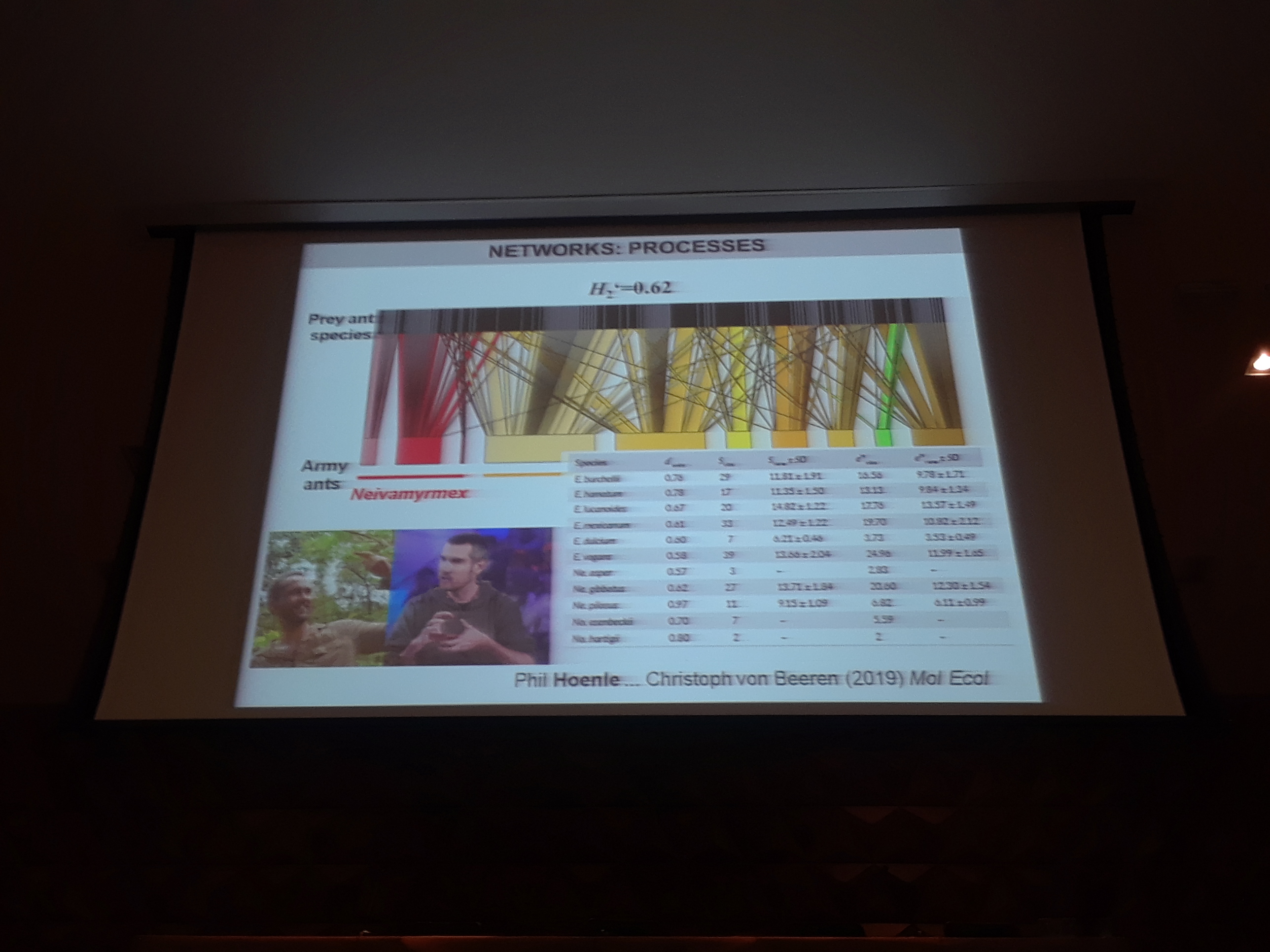
5:10pm: Moving from patterns to process, networks can also be used to investigate the causes of such patterns (or lack of patterns) by looking at traits that promote or inhibit interactions. Which traits structure each link in the network? Using similar null-model based approach, you can assess if some traits are overrepresented or underrepresented relative to the null hypothesis. In many studies over several years, Dr. Blüthgen and colleagues have found that olfactory signals structure flower visitor networks (including ants but also many other invertebrates like crickets and hunting spiders! Have floral scents evolved as a defensive trait (against ants perhaps?). Went to Hawaii, where there are no ants! And… in fact, they found that endemic and native plants are disproportionately visited by ants, which suck up their nectar.


4:59pm: Through an experiment in southern China, they found that plant-Hemiptera specialization higher than Hemiptera-ant specialization, both of which decrease with increasing tree species richness. Also, in different studies, Dr. Blüthgen and colleagues find variable results on the association between specialization and latitude across taxa.
4:52pm: Coffee is refreshing, but not as refreshing as ants. Thankfully, we have one final talk today, by keynote speaker Dr. Nico Blüthgen titled “Ants on plants: untangling the networks with natural history and chemistry”. Fair warning: This talk will include some non-ant insects (!). Broad question: How do ecosystems change, especially with respect to species interactions and ecosystem functioning? Because this works includes ample use of networks, Dr. Blüthgen first discusses some pitfalls to network use and analysis. There is a problem because the more observations you have, the more links you have – so, for example, it can be hard to know if a species is specialized or just rare, based on number of links (or interaction strength or diversity, or nestedness, or conductance) in a network. One potential better way to use networks is assessing patterns (e.g. comparing a null model to a real network). By creating a model that is independent of number of species or total abundance, you can directly assess specialization hypotheses. More on this study can be found in Blüthgen et al. (2007, Current Biology).



4:00-4:30pm: Coffee break…
3:42pm: For the final speaker of this panel, we have Dr. Phil Ward on “Homoplasy and heterogeneity in ant evolution: samples and insights from the ant tribe Camponotini”. Dr. Ward begins by introducing the Camponotini tribe, which is of course widespread and diverse in terms of both species (~2,500+ species) and morphology. This group has also seen significant taxonomic revisions in recent years through phylogenetic and phylogenomic research that reveal quite a bit of homoplasy and also differential divergence from the original body plan. Also, all camponotines harbor a unique, intracellular, vertically inherited bacterial symbiont Blochmannia, located in the ant ovaries and in specialized cells of the ant midgut. After this introduction, Dr. Ward announces a new research project “CAnBE”: Camponotini Ants and their Bacterial Endosymbionts, with the goal of inferring the evolutionary history of camponotine ants with UCE data, analyze genome evolution in Blochmannia across representative camponotine clades, and also evaluate the relationship between the phenotype and gene content of the symbiont. A preliminary phylogeny includes 1844 UCE loci in a 90% taxa complete matrix, and recovers the relationships found in previous work. One finding here is fairly high biogeographical fidelity in the main Camponotus clade, and a single colonization of the Neotropical region by camponotine ants roughly 25 million years ago suggesting convergent evolution of traits between Neotropical and other region camponotines. It is possible that Blochmannia may be linked to coevolution and possibly the success in this group.

3:18pm: We wouldn’t blame you for a bit of a case of whiplash, because now we’re returning to social parasites – with Dr. Rachelle Adams. Dr. Adams’ focus today is on the stages of successful exploitation (broken down into “encounter” and “exploitation” phases). This work uses a host-guest system where the host is a fungus-farmer (and the parasite species is in the genus Megalomyrmex). Interestingly, this system has two parasites (a more lethal raider ant species in Gnamptogenys) that competes with the Megalomyrmex parasite. The first challenge the parasite must face is… finding the habitat location of the host! Dr. Adams and colleagues investigate infiltration/integration hypotheses (chemical mimicry [of the host], chemical insignificance, and chemical weaponry [e.g. behavior disrupter]). Using chemical profile analyses, they detected a distinct chemical profile with very few hydrocarbons in the cuticle of Megalomyrmex, supporting the chemical insignificance hypothesis. But in looking at the entire profile, they also find evidence of venom alkaloids that support the chemical weaponry hypothesis. Taking a step back, Dr. Adams spoke more about Megalomyrmex behavior, including “gaster flagging” that looks like a fancy dance move. In a test relating to behavior in these parasites, they conducted 94 trials, with aggressive interactions found in a little over 50% of these trials. Neat social parasite behaviors were found, including inducing submission in the host, and also repelling host aggressive behavior with presentation of chemicals on the gaster via gaster flagging, which occurred in a majority of aggressive interactions. In the future, Dr. Adams plans to investigate, with colleagues, habitat selection and location while probing exploitation further as well.


2:53pm: Now we’re back with Dr. Andrea Lucky again! This time, she is speaking on “Tools for studying ant invasion biology: population genetics and phylogenomics meet applied entomology”. We live in the Anthropocene: “I might rename this ‘The Myrmecocene”, and where humans go, ants follow. The world has a lot of ant species, but only ~1% are exotic; however, 25% of Florida ant species are exotic. Can we predict the next global invaders? Can we prevent the next global invaders? Here, Dr. Lucky will discuss work in Nylanderia, aimed at predicting future invaders, determining invasive origins, and testing species boundaries. Before getting to this work, she notes that taxonomic accessibility and systematics is critical to the success of invasive species management, and as a field myrmecology has a lot of great resources. Now, for such a hard group like Nylanderia, the UCE phylogenomics approach should assist in resolving deep and shallow relationships! Goals: Identify lineages, reconstruct UCE-based phylogeny of the genus, determine age and origin of major clades, and evaluate species boundaries in six widespread species (three pairs). Dr. Lucky and colleagues found five independent origins of the globetrotting species, and four major clades based on biogeography with Indomalayan origin of 44.8 million years ago. For differentiating the six widespread species, found that only males could distinguish N. fulva and N. pubens morphologically. For N. guatemalensis and N. steinheili, these two come out monophyletic and thus differentiated… mostly, because a couple other species popped into the group. And in N. bourbonica and N. vaga, they also found differentiated clades. But, these species pairs require new species descriptions and new morphological characters – back too the taxonomic impediment! In closing, Dr. Lucky plugged the Invasive Ant Boot Camp (June 22-24, 2020)!
2:31pm: Hopping over to ICB Auditório Cerrado, Dr. Christian Rabeling presents on “Sympatric versus allopatric speciation: exploring the origins and evolution of social parasitism in ants”. There is a high diversity of social parasite species in ants, with around 350 known species engaging in the behavior (including undescribed species) with around 100 independent evolutionary origins known. But how did social parasites originate? Two key hypotheses: Interspecific and intraspecific (allopatric vs. sympatric speciation). Dr. Rabeling and colleagues investigated if there is evidence for sympatric speciation – and found that the parasitic species is phylogenetically nested within the host species, likely due to divergence being extremely recent coupled with incomplete lineage sorting. And, of course, sister pairs of host-parasite species always have sympatric (overlapping) distributions owing to the nature of the interactions! The host and parasite do not overlap during reproductive events, either. Thus, the evidence is consistent with a sympatric speciation hypothesis. Next, they inferred novel phylogenies of parasite-host groups, and found different results in different parts of the phylogeny – sometimes finding sister pairs (or nested pairs), and sometimes finding parasite species phylogenetically distinct from the host species. Overall, patterns are consistent with both inter- and intra-specific hypotheses. Dr. Rabeling also briefly discussed a project with social parasitism in Formica ants, including a phylogeny in a manuscript under revision – and the genus is only 20-30 million years old. Dulosis (a type of brood-theft-based parasitism) arose once, and dependent colony foundation seems to be a prerequisite for the evolution of dulosis and inquilinism.


2:09pm: Dr. Vieira Costa reminds us that ants exhibit a high diversity of functional roles in interacting with ants (e.g. seed dispersal), and many drivers of ant behavior (e.g. resource quality). Some questions: Are plants that offer more nutritious rewards to ants better defended in return? Do plant rewards type mediate the outcome of interactions? What about impacts on space/distribution? Their work is in campo rupestre in Serra do Cipó – fitting for the name of this meeting room! They utilized Vochysia elliptica as the mutualistic plant species, and manipulative experiments to vary nectar resource quality. Additionally, they conducted some community ecology work on 5 ant-plant interaction types and implemented a network analysis approach. Work at this site continued for two years to also better understand the system of interactions assembly in space. In the V. elliptica system, they discovered that recruitment rate, encounter rate, attack rate, and exclusion success rate were all significantly higher as resource quality increased – the same ant species changes their behavior in response to resource quality. For the second study, a complex network was presented, with 78 plant species, 30 species, and 800 interactions. Foraging strategies and aggressiveness are likely driving interaction assembly. Across space, Dr. Viera Costa and colleagues found that only five species performed 50% of the interactions across all sites, yet those species also had heterogenous distribution. A possible dominance hierarchy drives this structuring by the generalist species. Overall: Plant rewards quality really does impact ant behavior and interaction outcomes at distinct scales! See Pacelle et al. (2018, Biotropica), Costa et al. (2016, Plos One), and Poisot et al. (2012) for more on these studies.

2:00pm: After a pleasant, relaxing lunch break, we’re back in Auditório Campo Rupestre to hear Dr. Fernanda Vieira Costa speak on how “Plant rewards quality modulate ant behavior and the outcome of ant-plant interactions”. First, we have a brief introduction by panel organizer Dr. Laura C. Leal on the goals of this panel, which is to create a space to discuss mutualisms at different scales. By coincidence, all three talks today are on communities in Brazilian dry habitats (caatinga and ferraro)!
12:00-2:00pm: Almoço delicioso!
12:00pm: We’ve ran back to the main auditorium for Dr. Andrea Lucky’s presentation titled “Tools for Studying Ant Invasion Biology-Population Genetics and Phylogenomics Meet Applied Entomology” [Note: The actual talk name differed from that listed in the program – pending update], and arrived a bit late. But not too late to hear about the amazing citizen science projects led by Dr. Lucky. One finding, looking at three major U.S. cities, was that the most commonly collected exotic ant is Tetramorium immigrans, and even the most commonly-collected ant period in one city. Also, two kids, in backyard collections, documented this species in two states where it had not been documented before. Another project focused on species identity (are all the populations of T. immigrans actually one species?), reproductive biology (are they supercolonial?), genetic variation, and dispersal history. They found that most certainly all populations across the U.S. are part of one species based on extremely similar DNA, and that they are in fact not supercolonial either (e.g. no evidence of inbreeding). Furthermore, the U.S. population is not geographically structured (even with a minor amount of variation) – probably maintaining low genetic diversity due to hitching a ride on human transportation! Lastly, Dr. Lucky and colleagues found significant differentiation between USA and European populations (the U.S. population is a subset of the European population, suggesting a single genetic lineage (though not necessarily a single immigration event) colonized the U.S.



11:55am: Dr. Ramalho ended her talk promoting an interesting survey she is conducting (with colleagues) on parenting in myrmecology, towards a more diverse and inclusive scientific field. Stay tuned for more about this, and please answer the survey if you receive it!

11:45am: Returning to tiny things inside tiny things, Dr. Manuela De Oliveira Ramalho is up next to speak about her work on “What Can Different Biomes and Monocultures Tell Us About the Bacterial Community of Atta sexdens (Linnaeus, 1758)?” She begins with a discussion of the economic importance of A. sexdens to Brazil, as these leaf-cutter ants are considered pests to crops in, for example, the São Paulo region. Beyond this economic importance, this species is also part of a multi-species interaction, both with its fungal crop but also with bacterial symbionts. Dr. Ramalho notes that there has been an explosion of papers on bacterial symbionts in recent years (> 1,500), but only about 70 on ants to date. In this study, they look at if and how habitat and geography impact the A. sexdens microbiome. 75 colonies were collected across São Paulo state, and a 16S rRNA approach was used. Dr. Ramalho and colleagues detected a diverse community, including bacteria associated with citrus, that was also associated with geography.



11:30am: Heading to agricultural systems, we now have Dr. Cléa Dos Santos Ferreira on “The ‘Ant-Garden’ of the Dolichoderine Azteca paresis bondari as a Biodiversity Microcosm in Cocoa Plantations of South-East Bahia, Brazil”. [NOTE: This talk was presented in Portuguese, and thus the reporting for this talk is based on information gleaned from the English-language slides only]. In this system, there is an epiphyte-ant interaction, with advantages of epiphyte seed dispersal by the ants, and nesting cavities provided by the plants for the ants. Three ant-gardens were collected in an agroforested farm, ant activity was reduced by cooling, and they screened the ant-gardens for species in the laboratory. They found a number of species spanning multiple insect orders in the ant-gardens, including many larvae of Ceratopogonidae midges, the flies that include the main cacao tree pollinators (Forcipomyia spp.). Further work should focus on the diversity of interactions between the ants and these invertebrates, and also on the possibility that some parasites and predators (or commensals) use the ant or nest deterrence to shelter their brood development.

11:15am: Moving on to mutualisms, here’s Inácio José De Melo Teles E Gomes speaking about “An Integrative Approach on Mechanisms Underlying Benefits That Ants Provide to Plants: A Long-Term Manipulative Experiment on the Cercopia-Azteca System”. Their core question: Do plants grow faster as a result of benefits bought by ant colonization? To figure this out, they planted 200 seedlings and 54 months of monitoring plant growth for two groups (colonized by ants vs. uncolonized by ants). Plant height growth and number of leaves were both higher in colonized relative to uncolonized plants. Also found 30 pathogens (fungi) in uncolonized group, and only 1 in the colonized group, suggesting that ants are also providing anti-fungal benefits. Nitrogen increased with time since colonized, and no association in control group with time. Also, naturally-produced plant chemical and physical defenses decreased with time since colonization, suggesting plasticity in plant defenses depending on whether or not they are colonized by their mutualist ants.
11:00am: Trap-jaw ants are in the house, with a talk by Cândida Anitta Pereira Rodrigues on “Signs of Subcastes in the Worker Caste of Odontomachus chelifer“. Species in the subfamily Ponerinae generally rarely exhibits polymorphism (only known in Megaponera and Paraponera). However here, they noticed some qualitative evidence of a major and minor worker in O. chelifer, and thus tested if there is morphometric distinctions between major and minor workers. Second, they assessed behavior (31 behavioral traits), and found differences in several behaviors. Third, they evaluated if the cuticular chemical profile differed between the two groups of workers, and found no significant difference. Overall, given the morphological and behavior differences, it seems that Odontomachus may now join Megaponera and Paraponera in the ranks of Ponerine genera with at least one species with worker subcastes!
10:45am: We had to quickly move to the Auditório Campo Rupestre and arrived a bit late to Dr. Robert Fagundes’ talk on “Hierarchy of Interaction Dominance in Ant-Plant Mutualism: A network Approach”. They produced an interaction matrix from 200 hours of ant worker interactions recorded and analyzed, and found a clear dominance hierarchy with a Camponotus (carpenter ant) species at the top. And the sugar concentration of nectar provided by plants promoted increased hierarchization.


10:38am: To investigate if Pheidole has a core microbiota, a microbiota related to seed harvesting, or a microbiota related to geography, as well as explanatory factors related to phylogeny, Dr. Martins collected samples from around the world for 16S rRNA amplicon sequencing. Found a stable core microbiota, and in particular Proteobacteria and Firmicutes, as well as Wolbachia. Also detected Ralstonia – this is associated only with seed-harvesting species! Need further work to determine if these bacteria provide some function(s) related to seed-harvesting. Food resource and geographical location also associated with bacterial community. In concluding thoughts, Dr. Martins also promoted their instagram account promoting women in science, @mulheresnacienciaufpi!


10:31am: Up next, Dr. Cintia Martins is asking, “Does a seed harvesting diet, host phylogeny and geographical location influence the bacterial community of Pheidole ants?”
10:25am: Now time for the oral presentations! We’re in the main auditorium, where Cristian Luan Klunk is presenting on “Environmental resistance evolution in ants: effects of body size, cuticular brightness, and nesting layer”. For this study, they measured cuticular brightness and nesting layer (based on observations and the literature), and phylogeny was inferred, and tested effects of heat stress and cold stress resistance, as well as desiccation resistance. For heat stress, a positive association between body size and resistance was detected, while a negative association between cuticular brightness and heat resistance was detected. Brighter ants and larger ants performed better under cold stress. For desiccation stress resistance, body size exhibit a positive association and brightness a negative association. For nesting, only an association with desiccation stress was detected, with the ground nesting ants performing better, contrary to expectations (if we heard this result correctly!).
10:00am: Talk summary and concluding thoughts about museum collections and inclusion and diversity in science, captured in a video! Apologies for the shaking video…
9:53am: Now asking a surprising question. Does gut bacteria help ants produce stronger cuticle for their exoskeletons? Four studies done to date, all in beetles, found that when antibiotics given to beetles, cuticle becomes pale and thin. But which cuticular components are impacted? Are other pathways involved than the one so far established? Are mechanisms conserved across species? The manuscript on this work (Duplais and Moreau) is in preparation, so we will leave some details unstated here. But overall, bacteria appear to be quite important for ant cuticle production in various ways, with some differences from work in beetles!
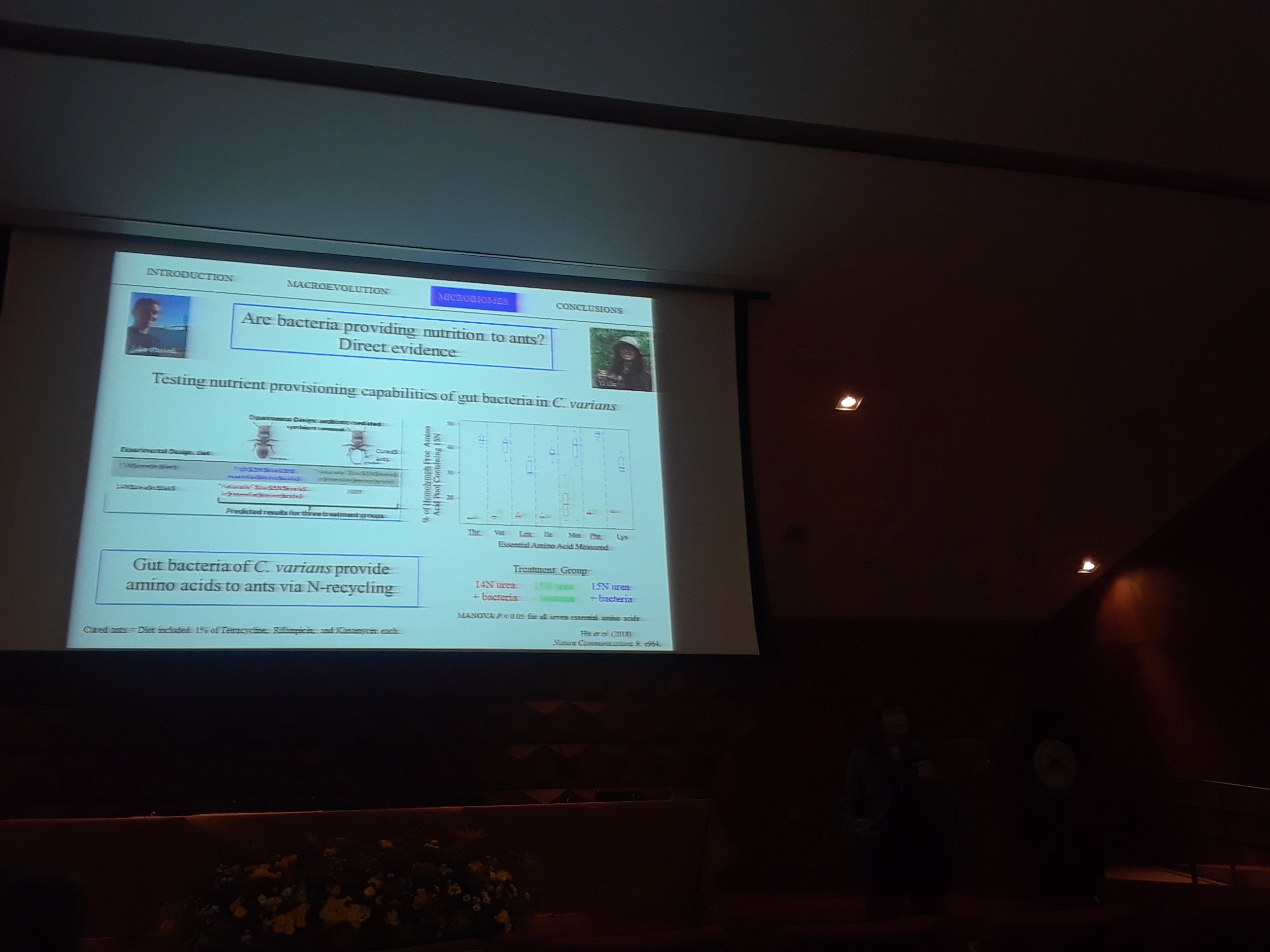
9:45am: Time for some things even tinier than ants. In other words, bacteria. “Ants provide an excellent system to study the diversity, distribution, and influence of gut-associated bacteria”. Diet & nutrition, transmission, host-symbiont coevolution, and geography all of interest to Dr. Moreau. First work reported in Russell, Moreau et al. (2009, PNAS). If you feed low on trophic scale, you have particular bacteria in your gut (Rhizobiales). Five independent evolutions of associations with Rhizobiales in ants. Are bacteria providing nutrition in ants? Indirect evidence, reported in Kautz et al. (2013, Applied Environmental Microbiology). Are the bacteria assisting Cephalotes to digest pollen? Pollen has a tough outer coating! Increase in Rhizobiales detected when pollen re-introduced to diet. But we now also have more direct evidence as well: Metagenomic sequencing reveals that core bacteria from turtle ant guts synthesize essential amino acids. Check out Hu et al. (2018, Nature Communications) for more on this interesting work.
9:30am: Moving on to… genome evolution in ant-plant mutualists! Dr. Moreau referencing the Red Queen Hypothesis now – “it takes all the running you can do, to keep in the same place”. Examples of parasitic relationships following this pattern: a parasitic organism must continue to evolve as the partner evolves, to maintain parasitism. Now it’s counterpart, the Red King Hypothesis, where evolving more slowly is favored. We predict seeing this in mutualism. Let’s test this! Enter Pseudomyrmex plant-ants. There are three independent evolutions (i.e. convergent evolution) of obligate mutualism, which is perfect as they are natural replicates well-suited to testing predictions. Generated reference genome (at a time when no ant genome had yet been produced!) and mapped genomes of other species in analysis to this reference genome. Two most important findings: (1) strong convergent positive selection on genes related to neurogenesis, and (2) elevated rates of genome evolution in mutualists. Difference in rates not due to differences in generation time (mutualists actually have longer generation times) or population size (dN/dS ratios do not differ between mutualists and generalists), but may be due to relaxed selection and/or elevated mutation rate. Overall, results contradict expectations of the Red King Hypothesis, and suggest that the Red Queen Hypothesis should fit all types of obligate symbioses! See Rubin and Moreau (2016, Nature Communications) for more.
9:16am: To answer this question, Dr. Moreau, Matt Nelsen, and colleagues inferred a phylogeny of almost 11,000 vascular plant species, and a phylogeny of 1,730 ant species. Wow! Extrafloral nectaries evolved early, domatia evolved later. And ants go from predatory feeding strategy to more general diet to herbivory – no jumps from predator to herbivorous, and no reversals. Once an herbivorous ant, always an herbivorous ant! Incremental sequence of evolutionary events led to full ant reliance on plants, and diversification rates of plant-associated ant lineages are not enhanced. See Nelsen, Ree & Moreau (2018, PNAS) for the scientific article on this research.
9:12am: “No organism acts in isolation”. Dr. Moreau presenting research on coevolutionary interactions between ants and plants, and ants and bacterial communities. Are plant and ant evolution linked?
9:09am: Time to learn about macroevolution and microbes and, obviously, das formigas!

8:48am: We got here early, and the main auditorium is being prepared for Dr. Moreau’s presentation. Oh, the ant-icipation!!!

8:20am: We’re beginning our live coverage on the second full day of the Symposium, which promises plenary talks by keynote speakers Dr. Corrie Moreau, of Cornell University, and Dr. Nico Blüthgen of Technische Universität Darmstadt, as well as a number of exciting oral presentations. We will provide post hoc coverage of the first day of the conference based on notes taken on that day – stay tuned! Up next: Dr. Corrie Moreau on “Piecing Together the Puzzle to Understand the Evolution of the Ants: Macroevolution to Microbiomes” in Auditório Nobre, CAD1.

Leave a comment